NOTE: This is from the biological report on the status of Atlantic Salmon - see Table of Contents and News Release for additional information.
7.5 OTHER NATURAL OR MANMADE FACTORS AFFECTING ITS CONTINUED EXISTENCE
7.5.1 Aquaculture: Atlantic Salmon Farming
7.5.1.1 Production and Location of Sites
The development and expansion of Atlantic salmon aquaculture has occurred in the North Atlantic since the early 1970's. Worldwide production of farmed Atlantic salmon in 1998 was 710,342 tons, over 295 times the nominal catch of Atlantic salmon in the North Atlantic (ICES WG 1998). Production in the North Atlantic area in 1998 was 538,011 tons, of this Norway produced 64% and Scotland produced 21% (ICES WG 1998). In the United States, Atlantic salmon aquaculture occurs predominantly in the states of Washington and Maine. In the Gulf of Maine, the industry has grown rapidly in the last decade. Annual Atlantic salmon aquaculture production in Maine increased from an estimated 10 mt in 1984, to 454 mt in 1988, to 12,250 mt in 1997 (Honey et al. 1993; Baum 1998).The Maine Atlantic salmon aquaculture industry is currently composed of 12 companies, at 33 sites with a total of 773 cages covering 800 leased acres of water. Farms are concentrated in Cobscook Bay near Eastport, Maine but are located as far south as the Sheepscot River, although that site currently does not grow Atlantic salmon. The industry in Canada, just across the border, is approximately twice the size of the Maine industry.
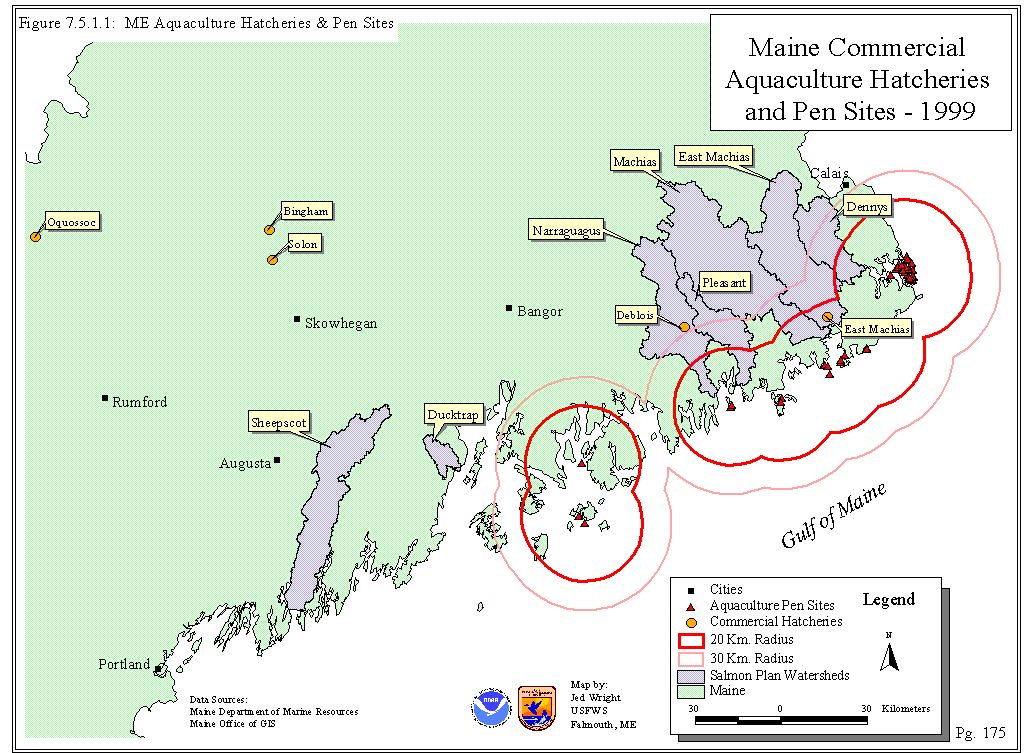
Five freshwater hatcheries in the United States provide smolt to the sea cages and produce up to four million smolt per year. Locations of cages and freshwater hatcheries are displayed in Figure 7.5.1.1. It takes approximately 18 months to grow a market-sized fish and annual production in Maine is about six million salmon or 12,000 mt (Baum 1998). Three broodstock lines are used for farm production. The lines include fish from the Penobscot River, St. John River, and an industry strain from Scotland. The Scottish strain was imported into the U.S. in the early 1990s and is composed primarily of Norwegian strains, frequently referred to as Landcatch. In recent years, milt of Norwegian origin has been imported by the industry from Iceland (DFO 1999). It is estimated that there is European genetic influence in approximately 30 to 50% of the production fish in Maine (Baum 1998)
Figure 7.5.1.1: Commercial Aquaculture Hatcheries and Pen Sites
{kind=link}
7.5.1.2 Threats to Wild Salmon
Atlantic salmon that escape from farms and hatcheries pose a threat to native Atlantic salmon populations in coastal Maine rivers. Escapement and resultant interactions with native stocks are expected to increase given the continued operation of farms and growth of the industry under current practices. There is a potential for escaped farmed salmon to disrupt redds of wild salmon, compete with wild salmon for food and habitat, interbreed with wild salmon, transfer disease or parasites to wild salmon and/or degrade benthic habitat (Clifford 1997; Youngson et al. 1993; Webb et al. 1993; Windsor and Hutchinson 1990; Saunders 1991). A comparison study in Canada revealed that survival of wild post-smolts moving from Passamaquoddy Bay to the Bay of Fundy was inversely related to the density of cages (DFO 1999). Finally, there has been recent concern over potential interactions when wild adult salmon migrate past closely spaced cages, creating the potential for behavioral interactions, disease transfer or interactions with predators (DFO 1999; Crozier 1993; Skaala and Hindar 1997; Carr et al. 1997; Lura and Saegrov 1991).Farm raised Atlantic salmon have been documented to exist in the wild because of escapement and release from salmon aquaculture facilities (ICES-NASWG 1994; NASCO 1993; Bergan et al. 1991; Lura and Saegrov 1991; Hansen et al. 1993; Skaala and Hindar 1997; Stokesbury and Lacroix 1997). In Norway, the number of salmon that escape from sea cages is thought to be greater than the number of salmon in the wild (Gausen and Moen 1991). Monitoring on 30 rivers in Norway in 1997 documented farmed fish in 26 rivers. Between 1992 and 1997, the total stock size of wild and hatchery origin adult Atlantic salmon returning to the Canadian Maritimes was between 115,000 and 229,000. During that same time, the number of salmon in net pens in the Bay of Fundy exceeded fifteen million. It is estimated that 25-40% of the fish in the North Atlantic Ocean is of aquaculture origin (Jonsson 1997). In Atlantic Canada, most Atlantic salmon aquaculture occurs in the lower portion of the Bay of Fundy. There are an estimated 60 aquaculture facilities (farms) in the area, and salmon that have escaped from farms have been documented in New Brunswick rivers (ICES-NASWG 1994).
Reports of large-scale escapes are rare; however, in 1994 there was one reported escape in New Brunswick of between 20,000 and 40,000 fish which was equal to the total estimated wild returns to Nova Scotia and New Brunswick that same year. Since the aquaculture industry began in 1979 in the Maritimes, escapees have been documented in 14 rivers in New Brunswick and Nova Scotia (DFO 1999). The Magaguadavic River is monitored for interactions between wild and farmed fish in Canada. In at least two years, over 90% of the adult salmon entering the Magaguadavic River were of farmed origin. This data has been used to demonstrate the fact that the three aquaculture hatcheries in the watershed are leaking farmed juveniles. Emigrating smolts in 1996 were 51-67% farm-origin and those exiting the river in 1998 were 82% farm-origin (DFO 1999). Analysis of eggs taken from the Magaguadavic River in 1993 revealed that at least 20% of the redds were constructed by females of farm or cultured origin, and another 35% were of possible cultured origin (Carr et al. 1997).
Atlantic salmon that either escaped or were released from farms have been found in the St. Croix, Penobscot, Dennys, East Machias, and Narraguagus rivers in the U.S. (Baum 1991; USASAC 1996, 1997). In 1994 and 1997, escaped farmed fish represented 89% and 100% respectively, of the documented run for the Dennys River, and in 1995, 22% of the documented run for the Narraguagus River (Table 7.5.1.2). Escaped farmed salmon have also been documented as an incidental capture in the recreational fishery, or observed in additional rivers: Boyden, Hobart, and the Pennamaquan Rivers. The first aquaculture escapee in the State of Maine was documented in 1990 and the first sexually mature escapee was documented in 1996. Escaped farmed fish are of great concern in Maine because even at low numbers they can represent a substantial portion of the returns to some rivers. Also, populations at low levels are particularly vulnerable to genetic intrusion or other disturbance caused by escapees (DFO 1999, Hutchings 1991). Preliminary results from the 1999 wild smolt assessment project in the Pleasant River, Washington County, suggest that several outmigrating smolts were of hatchery origin based on fin condition (Kocik et al. 1999, unpublished data). Genetic and scale samples were collected to better determine the origin of these fish.
Table 7.5.1.2. Number and percentage of the total run of farmed salmon found in the Dennys and Narraguagus rivers, 1993-1997.
|
YEAR |
DENNYS |
NARRAGUAGUS |
||
|
Number |
% of Run |
Number |
% of Run |
|
|
1993 |
20 |
45% |
1 |
2% |
|
1994 |
42 |
89% |
0 |
0% |
|
1995 |
4 |
44% |
8 |
22% |
|
1996 |
21 |
68% |
0 |
0% |
|
1997 |
2 |
100% |
1 |
4% |
Because of selection in the hatchery or culture environment, farmed salmon are expected to be less fit for life in the wild. However, they may have competitive advantages at certain life stages (Gross 1999). Experimental tests of genetic divergence between farmed and wild salmon indicate that farming generates rapid genetic change as a result of both intentional and unintentional selection in culture and that those changes alter important fitness-related traits (McGinnity et al. 1997; Gross 1998). This change was identified as a threat to wild populations when cultured fish escape, and compete and breed with wild salmon (Hindar et al. 1991; Fleming and Einum 1997). Farmed salmon in Scandinavian countries have been documented to spawn successfully, but later in the season than wild salmon (Lura and Saegrov 1991; Jonsson et al. 1991), a factor that increases the potential for limiting the success of wild spawners through redd superimposition. Culture or artificial propagation selects for attributes that affect the behavior of farmed salmon (Hindar et al. 1991; Utter et al. 1993).
Study results were presented at the 1997 joint ICES/NASCO Symposium on Interactions between Salmon Culture and Wild Stocks of Atlantic salmon which demonstrated that spawning between wild and cultured fish would have negative consequences for the wild stock since the offspring of the cultured fish occupied juvenile habitat and displaced wild fish (CNL (97)27). Escapees in Europe provide evidence of spawning success and also of redd superimposition (DFO 1999). Mork (1991) characterized the potential permanent effect of one generation burst immigrations, resulting from large scale escapes from farms near spawning rivers, on the genetic differentiation among wild stocks. He reported that small Atlantic salmon populations may be most vulnerable to burst immigrations, and these events could be the most significant way in which farmed salmon affect the genetic structure of wild populations. Natural selection may be able to purge wild populations of maladaptive traits but may be less able to if the intrusions occur regularly year-after-year. Under this scenario, population fitness is likely to decrease as the selection from the artificial culture operation overrides wild selection (Fleming and Einum 1997; Hindar et al. 1991). Genetic interactions between wild and farmed fish can disrupt local adaptations, threaten stock viability and character, and lower recruitment (DFO 1999; Einum and Fleming 1997; Fleming and Einum 1997; Grant 1997; Saegrove et al. 1997).
A study was conducted in a natural spawning tributary of the Burrishoole River system in western Ireland to compare the performance of wild, farmed, and hybrid Atlantic salmon progeny. Survival of progeny of farmed fish to the smolt stage was significantly lower than that of wild salmon. The progeny of farmed fish, however, grew faster and displaced native fish downstream (McGinnity et al. 1997). This study demonstrated that both farmed fish and hybrids can survive in the wild. It also indicates that escaped farmed salmon can produce long term genetic changes in natural populations (McGinnity et al. 1997). The authors caution that repeated intrusions of escaped farmed salmon will depress smolt productivity in a cumulative fashion potentially creating an extinction vortex (McGinnity et al. 1997).
Initial studies in Ireland had focused on the potential for adults to escape from sea cage sites, locate and enter a wild river, and interbreed or otherwise interact with the wild population. However, recently the focus has been expanded to include concerns over escapes from freshwater hatcheries in Ireland on rivers with wild populations, and potential competition and interaction between these escapees and wild stock from early life stages to spawning adults. A relationship has been demonstrated between the reproductive success of cultured fish and the time the fish has lived in nature before reaching sexual maturity (Jonsson 1997). Consequently, escapees from freshwater hatcheries may pose a larger threat to wild populations than escapees from sea cage sites.
Experiments have been conducted in rearing triploid Atlantic salmon with the hope that the use of these sterile fish in commercial culture would eliminate concerns for genetic interaction with wild stock. While growth and survival in freshwater has been demonstrated to be comparable to diploid strains, mortality during the transition to marine cages has been higher and deformities among triploids remain a major concern (O'Flynn et al. 1997). A comprehensive evaluation of the use of sterile triploid Atlantic salmon was undertaken from 1994-1998 by the Marine Laboratory in Aberdeen. Performance trials in Ireland and Norway demonstrated that triploids grew similarly and survived as well as diploids in freshwater. In sea water, triploids grew similarly to diploids but suffered higher losses in half of the trials (Marine Laboratory Aberdeen 1998).
Given current aquaculture practices, the Services have opposed the use of reproductively viable European strains (pure and hybrid) of Atlantic salmon within North America. This opposition is based on genetic studies that demonstrate that there are significant differences between North American and European Atlantic salmon (King et al. 1999), and the advice from geneticists that interbreeding among genetically divergent populations negatively impacts natural populations (Utter 1993; Verspoor 1997; Youngson and Verspoor 1998). The introgression by non-North American Atlantic salmon stocks presents a substantial threat of disrupting the genetic integrity of North American stocks and threaten fitness through outbreeding depression.
Disease epizootics in wild Atlantic salmon populations are not common, but in culture operations where fish are artificially propagated under high densities, they can pose a significant threat (NASCO 1993; Saunders 1991). Diseases affecting Atlantic salmon reared in captivity include bacterial, parasitic, viral, fungal and nutritional diseases (Roberts 1993). The development of a disease epizootic results from an interaction between the host, environment and the disease agent. In farmed salmon, the occurrence of disease is generally due to the high densities at which fish are reared (Hastein and Lindstad 1991). Bacteria may be released to the environment during and after epizootic diseases and may survive and persist (Olafsen 1993; Egusa 1992). The occurrence and spread of infectious diseases increases due to the high densities at which farmed salmon are raised (Institute of Aquaculture 1988; Lura and Saegrov 1991; Hastein and Lindstad 1991; Mork 1991; NASCO 1993; Olafson 1993).
The disease interaction between wild and farmed salmon will likely occur through the water, fish, and other sources such as nets and fishing or handling gear. The transmission of diseases through water can take place over long distances, and transmission has been documented to occur over at least seven km (Hastein and Lindstad 1991). Incidences of disease transmission from Atlantic salmon farms to wild stocks are not well documented, but investigations are becoming more numerous. The greatest risk associated with diseases is the introduction of an exotic disease into a new area where local stocks have no innate resistance (McVicar 1997, DFO 1999). In Norway, over thirty populations of salmon have extirpated by the parasite Gyrodactylus salaris (Heggberget et al. 1993).
In response to the presence of Infectious Salmon Anemia (ISA) in Canadian aquaculture sites, the Maine aquaculture industry initiated a biosecurity audit to identify strengths and weaknesses in farm-level disease prevention procedures, to prevent the spread of ISA into Maine and to detect the potential emergence of ISA at the cage level. The Maine Aquaculture Association has also reported that emergency eradication procedures for exotic disease events have been adopted and updated state regulations governing the importation and movement of live fish and gametes are nearing completion.
As discussed above, ecological interactions between wild and cultured Atlantic salmon can occur at any life stage through competition for food and habitat, and during spawning. There is also the potential for cage sites to negatively impact the benthic habitat from discharge of wastes and excess food. In 1992, Maine initiated the Finfish Aquaculture Monitoring Program (FAMP) to evaluate the impact of marine cages on benthic habitat. The program is funded by a one cent per pound tax on production. While there have been isolated examples of degradation below particular sites, they were fallowed and found to recover quickly. There was also no evidence of negative impacts to wild salmon habitat, water quality or benthic organisms from wastes or the use of drugs and chemicals in the Maine aquaculture industry (Baum 1998).
In 1998, the Department of Fisheries and Oceans convened a workshop to assess the potential impacts of aquaculture on wild stock and to recommend ways to minimize the risks (DFO Maritimes Regional Habitat Status Report 99/1 E CITE FORMAT). Concern was heightened by the drastic drop in wild stocks in areas where aquaculture was conducted. For example, the spawning run in the Magaguadavic River decreased from 293 in 1992 to 31 in 1998. Egg deposition during that same time decreased from 80% to 2.3% of the conservation requirement. Workshop participants identified the following potential interactions between farmed and wild Atlantic stocks (relative risk to wild stocks is in parenthesis): competition in the ocean (unknown); disruption of migration by interference (unknown); redd superimposition (medium); competition for food and space in freshwater (high); habitat displacement in freshwater (high); altered predator-prey relations (high); disruption of migration by cages and hatcheries (unknown); and hybridization (medium-high) (DFO 1999).
The summary of this workshop recommended application of a precautionary approach for management of the Maritimes salmon stocks and their interaction with escaped farmed salmon. This approach was based on documentation of local interactions, evidence of genetic and behavioral interactions between escapees and wild salmon in Europe, predictions of a regional model assessing risk, and the potential for negative interactions. The following priority action items were identified to minimize the risk of interaction between wild and farmed salmon: improving containment; improving fish health management; upgrading the policy for introductions and transfers of fishes and improving related enforcement; enhancing education and training of aquaculture workers; maintaining wild stocks at or above their conservation requirements; continuing the use of local stocks as donors; and incorporating risk analysis into the process for siting hatcheries and farms (DFO 1999). A further recommendation was given that counting fences be used for all rivers with unique stocks in which escapees are found to be compromising a significant portion of the total returns (DFO 1999).
The Maine Aquaculture Association has developed and adopted a Code of Practice for the Responsible Containment of Farmed Atlantic Salmon in Maine Waters (October 1998). The Code focuses on net strength, equipment integrity and predator control measures. An annual four-point stress test is required for all nets greater than three years old. Cages are also to be marked with the name of the manufacturer, year produced, field assembler, date deployed, and an inventory control number. Written maintenance records are to be kept on site for all nets and cages. Additional components of the Code discuss moorings and freshwater hatchery containment. Operating procedures for fish transfers, predator control and storm preparation are specified to minimize the risk of escapement. However, the details of the reporting, monitoring and enforcement of the Code remain to be worked out at this point.
An additional action to be taken in an attempt to prevent opportunities for aquaculture escapees to interact with wild stock is the placement of weirs in wild salmon rivers. A trapping facility is already located on the Narraguagus River so migrating adults can be examined and obvious aquaculture escapees can be denied passage upstream to spawning sites. Currently, aquaculture escapees are identified primarily by condition of the fins and body shape. There has been discussion of pursuing a universal tag that could be placed on all aquaculture fish to ensure that identification at weir and other sorting sites would be absolute. Such a mark has not been developed at this time.
Permits have been secured for the placement of weirs on the Dennys and Pleasant rivers and the State plans to install these weirs by October 1999. Design work for a weir on the East Machias River is ongoing and construction is planned to begin this fall. The decision has been made to design a fish trap for the Machias River Gorge instead of a weir and the ASC and FWS are currently working on that design.
7.5.2 Natural Mortality in the Marine Environment
Natural mortality in the marine environment can be attributed to four general sources: predation, starvation, disease/parasites, and abiotic factors. While our knowledge of the marine ecology of Atlantic salmon has increased substantially in the past decade, we cannot partition total natural mortality into these categories. Consequently, investigations of natural mortality are currently based upon an examination of return rates or total marine survival. Estimates of total mortality can be made by relating either hatchery smolt stocking rates or estimates of wild smolt production to the return of adult spawners. This method integrates all natural mortality factors and, if applicable, fishing mortality. If smolts are enumerated near the marine environment, the return rate indexes only marine survival. If the smolts are enumerated as they are stocked into upstream reaches, then assessment of return rate will include outmigration mortality.Reported survival rates of Atlantic salmon during the marine phase range from 0-20%, based upon a review of 20 studies by Bley and Moring (1988). Our review of additional studies found that this range is realistic for Atlantic salmon survival and that most return rates fall in the lower quartile of this range (Reddin 1988; Ritter 1989; Scarnecchia et al. 1989). In the U.S., return rates have generally been less than 1.5% in the Penobscot River and even lower in the Connecticut and Merrimack Rivers. In fact, return rates for Connecticut and Merrimack River hatchery stocks average 12% and 27% of that of the St. John River in Canada, which is one of the closest Canadian rivers to the U.S. The average Penobscot River return rate is about 89% of the St. John River average. The Connecticut and Merrimack River Atlantic salmon return rates are very low compared to the rates observed in other predominantly 2SW populations. Wild stocks and stocks returning after one sea winter typically return at higher rates (Bley and Moring 1988). Lower return rates might be expected for U.S. stocks, which are primarily 2SW fish and have been the result of smolt releases for most of the restoration period. However, in a comparison to only the hatchery stocks of the St. John River, survival was still lower in the more southern U.S. systems (Porter and Ritter 1984).
Some investigators have suggested that Atlantic salmon stocks that utilize longer migration routes typically have lower marine survival (Bley and Moring 1988). The authors also note a north to south gradient of decreasing marine survival that is consistent with this hypothesis. This hypothesis also helps to explain the typically high survival seen in several of the northern (Icelandic and Irish) stocks of Atlantic salmon with limited migratory routes. As such, the lower return rates of U.S. stocks may be a result of their relatively long migrations. If this is the case, these lower return rates may simply reflect the geographic location of these stocks in the southern extent of the range of Atlantic salmon. It is important to note that there is also a north-south trend of decreasing smolt-ages. This trend results in higher freshwater productivity in the southern extent of Atlantic salmon range that would offset the higher marine mortality.
On an interannual basis, marine survival rates can be more variable than freshwater survival rates. Reddin (1988) evaluated the freshwater (egg to smolt) and marine (smolt to spawner) survival for seven cohorts of Atlantic salmon in West Arm Brook. He found that marine survival was typically higher (5.51%) than freshwater (1.67%). However, the variation in marine survival, as measured by the coefficient of variation, was nearly four times greater in the ocean (63%) than in the stream (14%). These results were partly confounded by the fact that these stocks are exploited at sea, albeit only lightly. However, unexploited Icelandic stocks had similar variation (62%) in marine survival (Scarnecchia 1984a; Scarnecchia et al. 1989). Thus, the production potential and population dynamics of Atlantic salmon may be determined by year-to-year variability of oceanic natural mortality as well as the average level of natural mortality in the marine environment.
The year-to-year variation in return rates of U.S. stocks is generally synchronous with other Atlantic salmon stocks although at lower absolute levels (Friedland et al. 1993). Recent return rates have been decreasing for several North American Atlantic salmon stocks. This suggests that while some factors distinct to the U.S. stocks may be causing low return rates, the general trend is being driven by a factor that occurs when the stocks are mixed. Friedland et al. (1993) documented a common pattern of return rates for five North American stocks, including the Penobscot River and Connecticut River stocks, suggesting that all of these stocks responded equally to variation in survival. This observation provides an alternate hypothesis to conventional thinking that the most significant natural mortality occurs in the river, estuary, and close to the river mouth (Larsson 1985; Wood 1987; Hvidsten and Lund 1988; Magnhagen 1988). The correlations between the survival rates suggest that an important cause of mortality may act upon the stocks when they are mixed and utilizing a shared habitat. Since North American Atlantic salmon are migrating from geographically distinct rivers to common ocean feeding grounds, the likelihood that their distributions will begin to overlap increases with the length of marine residency. Thus, autumn and winter are the most likely seasons when post-smolt survival is determined. Similar recruitment cohesion has been described in other Atlantic salmon stocks and salmonine species (Scarnecchia 1984a; Koslow et al. 1987; Cohen et al. 1991). This observation indicates that factors occurring in the North Atlantic, and particularly the Labrador Sea, may be important to the survival of many Atlantic salmon stocks.
Survival rates are likely to be a function of growth patterns. Friedland et al. (1993) found that the survival rate for the Penobscot River stock was correlated to a growth index defined by intercirculi spacing over the winter period, suggesting that the first winter at sea regulates annual recruitment. This agrees with the analyses of Reddin and Shearer (1987) and Ritter (1989). This growth index also provides insight into the relationship between mortality and growth. Friedland et al. (1993) found an association between growth and survival such that in years of poor growth, a greater proportion of the stock died. When growth was better, so was survival. This suggests that the functional relationship between growth and survival may not be a threshold phenomenon. If a threshold was necessary for survival, the sample of scales from Atlantic salmon returning to rivers would only be obtained from those fish above a critical length (Friedland et al. 1993).
Sea surface temperature (SST) may be an important feature of the marine environment that affects Atlantic salmon survival. Saunders (1986) and Reddin and Shearer (1987) found that SST influenced Atlantic salmon marine distribution. Atlantic salmon were common in 4o to 10o C waters, a temperature range thought to be ideal for growth (Saunders 1986). Scarnecchia (1984a and 1984b) showed that temperatures were related to Icelandic Atlantic salmon production for both 1SW and 2SW groups. Reddin and Shearer (1987) tested Dunbar and Thomson's (1979) hypothesis that sea temperatures and distributions influenced Atlantic salmon abundance in West Greenland. They found that below-normal surface temperatures in the Labrador Sea over the winter were responsible for low catches in West Greenland in 1983 and 1984. Because homewater catches for the same stocks that occurred in West Greenland were also low, they postulated that the low temperatures decreased overall production. Friedland et al. (1993) and Reddin et al. (1993) found that the pattern of stock production was related to the area of winter habitat available to North American post-smolts. The lack of a relationship for spring, summer, and autumn suggests that habitat during these seasons may not be limiting. As such, these researchers concluded that a significant proportion of the variation in recruitment was the result of post-smolt survival. The area of habitat could be related to intraspecific competition for space and food resources and to predation effects on post-smolts (Friedland et al. 1993). While these investigations have indicated the importance of SST to Atlantic salmon recruitment, the mechanisms responsible for reduced survival are still unknown. Mortality could arise from stress, starvation, predation, disease, and, perhaps, other unknown mechanisms. Further research needs to be undertaken to fully understand the processes involved
In summary, based upon recent research, major seasonal events influence post-smolt survival in Atlantic salmon. It appears that survival of the North American stock complex of Atlantic salmon is at least partly determined when they are concentrated during the winter months in the habitat formed at the mouth of the Labrador Sea and east of Greenland (Reddin and Shearer 1987; Friedland et al. 1993; Reddin et al. 1993). The habitat may limit total North American stock production through intraspecific competition during this period, but the post-smolt year is not growth-limiting to individual fish. Limitations result in annual variation in survival and growth that are correlated to SST. Until more direct observation on the marine ecology of post-smolts during winter can be made, researchers must assume that mortality is controlled by the interaction of growth and predation.
7.5.3 Artificial Propagation and Atlantic Salmon
In 1992, the ASRSC and the USFWS implemented a river-specific stocking program for Maine rivers. The program was initiated for the following two reasons: runs were declining in the seven rivers in the DPS and numerous studies indicated that restocking efforts are more successful when the donor population comes from the river to be stocked. This river-specific stocking policy is consistent with the goal of the Maine Atlantic salmon program to maximize production of wild smolts by restocking river specific stocks and emphasizing fry releases (Moring et al. 1995). The natural Atlantic salmon population in these seven rivers is very low and artificial propagation, by eliminating significant mortalities in vulnerable early lifestages, has potential to increase the number of fish in a short time period, thereby avoiding extinction. Collection of wild parr and captive rearing also acts as a multiplier of spawner numbers increasing effective population size. The ultimate goal is to reach the stage where stocking is no longer necessary on a continual basis. The Prelisting Recovery Plan (Baum et al. 1992) formalized the commitment to manage the Downeast rivers as genetically viable and distinct stocks.The Prelisting Recovery Plan included the recovery objective of using restocking efforts with river-specific fry as a means to stabilize or increase river populations in the seven rivers in the DPS. The immediate need for action was justified by the apparent fragile status of the existing wild Atlantic salmon stocks in Maine. Returns of Atlantic salmon to Maine rivers from 1970 to 1993 were analyzed and the percentage of the run that was of natural origin was calculated. Atlantic salmon determined to be natural, either could have spent their entire lives in the wild, truly wild fish, or could have been stocked in the river as fry. The average return rate was calculated for years in which no adults could have returned from fry stocking and this was termed the wild return. Secondly, the average return rate was calculated for years in which returns from fry stocking would be possible, and this was termed the natural return. This analysis could lead one to conclude that fry stocking would not aid in the recovery of these runs. However, fry stocked during those years originated out of the basin. When this same analysis was conducted for the Penobscot River the opposite trend was observed. The percentage of the Atlantic salmon run of natural origin was higher during years influenced by previous fry stocking. The Penobscot River, unlike the rivers in the DPS, has had a predominately river-specific stocking program. Consequently, the Services have reason to believe that the DPS rivers will benefit from river-specific fry stocking.
The BRT has determined that artificial propagation, with river-specific stocks in conservation hatchery mode, is an appropriate tool for rebuilding naturally reproducing Atlantic salmon populations in the DPS. The purpose of the ESA is to conserve endangered species or threatened species and the ecosystems upon which they depend. Consequently, the goal is to restore the species to a point where it is self-sustaining without the aid of hatcheries. The Act defines conservation as the following: "the use of all methods and procedures which are necessary to bring any endangered species or threatened species to the point at which the measures provided pursuant to this Act are no longer necessary." The definition goes on to state that one such method is propagation. The NMFS has analyzed the role of artificial propagation under the ESA in relation to Pacific salmon (Hard et al. 1992). That analysis cautions that artificial propagation, as a conservation tool, must be conducted in a manner so as to maintain the distinctness of the species unit. The current policy, to adhere to river specific stocking in the DPS rivers as long as a minimum effective population size is maintained, is consistent with maintaining distinctness of these stocks (Waples 1990).
The NMFS and FWS have issued a draft policy addressing the role of controlled propagation in the conservation and recovery of species listed as endangered or threatened under the ESA (61 FR 4716). While the Gulf of Maine DPS of Atlantic salmon is currently listed only as a candidate species by the NMFS, and not as threatened or endangered, the river-specific Atlantic salmon program was still evaluated to determine if it was consistent with the policy. The proposed policy sanctions the controlled propagation of listed species when recommended in an approved recovery plan and supported by an approved genetics management plan. The Atlantic salmon river specific rearing program was an essential component of the pre-listing recovery plan drafted for the species and is operated according to the broodstock collection and spawning protocols designed to maintain the genetic diversity of the DPS. These protocols were adopted by the TAC in 1997 (Beland et al. 1997).
In determining whether or not the hatchery fish should be considered part of the listable entity on the west coast, the NMFS asked the question of whether there were appreciable differences between the hatchery and wild fish in characteristics believed to have a genetic basis (Hard et al. 1992). The decision to use river-specific stock is predicated on the belief that recovery has the greatest likelihood of success if the stock in the hatchery has the same genetic makeup as the wild stock in the river. Consequently, it is the Team's position that the hatchery-held river-specific stocks of Atlantic salmon from the DPS rivers of Maine should be included in the DPS. The Atlantic salmon from these rivers currently being held for artificial propagation, as well as their progeny, should be included in the DPS under review. It is important to note that listing decisions and recovery are based on the viability of the stock in the natural environment. Artificial propagation is not to be used as a substitute action for addressing the factors threatening the survival of the species in its natural environment but rather as a tool to prevent extinction while other threats are addressed, thereby allowing natural recovery to occur.
7.5.4 Other
Based on concern over low returns in 1997, the Department of Fisheries and Oceans (DFO) in Canada convened a workshop to examine trends in abundance of Atlantic salmon and factors that could have contributed to low returns (Atkinson et al. 1998). The following potential factors affecting sea survival of salmon were identified: (1) environmental conditions such as temperature and salinity; (2) removals in legal and illegal fisheries; (3) predation by cod, seals, seabirds, etc.; (4) diseases or parasites; or (5) a suite of other factors including changes in biological characteristics of stocks and effects of escaped aquaculture salmon. Freshwater production and abundance of smolts was not found to be the cause of the low returns in 1997. Commercial catches and/or bycatches in inshore and offshore fisheries were also eliminated as potential causes of low returns in 1997. Likewise, the availability of forage, ocean climate, pollution, disease, poaching, age at maturity and age of spawners were eliminated as causes for low returns in 1997. Predation by birds, seals, and cod was the remaining factor that could not be discounted as a potential cause of the low returns. The BRT believes that these analyses are equally true for Atlantic salmon of the Gulf of Maine DPS since marine ecology issues are similar and marine habitat overlaps substantially with Canadian stocks.7.5.5 Summary of Other Factors
Farm-raised Atlantic salmon can escape from both sea cages and freshwater hatcheries and enter rivers within the Gulf of Maine DPS as sexually mature adults. Available genetic data and visual observations indicate that aquaculture escapees may have successfully interbred with wild Atlantic salmon. Under current aquaculture practices, this problem will persist because the escapement of aquaculture salmon, and their interactions with wild stocks, is expected to increase with the continued operation and growth of the industry in the State of Maine.There is a significant potential for escaped aquaculture salmon to disrupt redds of wild salmon, compete with wild salmon for food and habitat, interbreed with wild salmon, and transfer disease or parasites to wild salmon. Comprehensive protective solutions to minimize the threat of interactions between wild and aquaculture salmon have not been implemented. The threat of these interactions is considered critical, given the fact that wild salmon stocks within the DPS are at low abundance levels, and are particularly vulnerable to genetic intrusion or other disturbances caused by escaped aquaculture salmon.
Studies have characterized the potential permanent effect of salmon escapes from farms on the genetic differentiation among wild stocks. Small Atlantic salmon populations, similar to those found in the DPS, are the most vulnerable to immigrations from aquaculture escapees. These immigration events may be one of the most significant ways in which aquaculture salmon affect the genetic structure of wild populations. While natural selection may be able to purge wild populations of maladaptive genetic traits, regularly occurring intrusions of aquaculture salmon into wild populations make this considerably less likely. Thus, scientific information indicates that interactions between wild and aquaculture salmon in the DPS may lead to decreased numbers of wild Atlantic salmon, and in the extreme, to extinction of the wild stock.
Marine survival rates continue to be low for U.S. stocks of Atlantic salmon, and the subsequent low abundance of salmon impedes recovery of the DPS. Scientists have attributed natural mortality in the marine environment to sources that include stress, predation, starvation, disease and parasites, and abiotic factors. In addition, scientific studies indicate that year-to-year variation in return rates of U.S. salmon stocks is generally synchronous with other North Atlantic stocks. This aspect suggests that the trend in return rates is the result of factors that occur when the stocks are mixed in the North Atlantic, particularly the Labrador Sea. Scientists have concluded that a significant proportion of the variation in recruitment or return rate is attributed to post-smolt survival. However, the mechanisms responsible for reduced post-smolt survival are still not well understood.
There is agreement among scientists that additional research should be conducted to better understand the
processes
or mechanisms responsible for reduced post-smolt survival, and such research is being pursued. There is also
consensus that action necessary to ensure survival of salmon stocks and to rebuild stocks within the DPS includes
artificial propagation. The Atlantic salmon river-specific recovery program has been identified as an essential
component of the strategy to rebuild salmon stocks in the DPS. This program has been designed and implemented
to maintain the genetic diversity and distinctness of the DPS. Because the abundance of wild salmon stocks in the
DPS is so very low, artificial propagation, through a river-specific stocking program is considered an important
tool
to maximize the production of natural smolts with genetic traits necessary for survival of the species.