NOTE: This is from the biological report on the status of Atlantic Salmon - see Table of Contents and News Release for additional information.
SECTION 5: CONSIDERATION AS A "SPECIES" UNDER THE ESA
5.1 DISTINCT POPULATION SEGMENT ANALYSIS
The ESA considers "any subspecies of fish or wildlife or plants, and any distinct population segment [DPS] of any species of vertebrate fish or wildlife that interbreeds when mature" to be a species. One of the purposes of this definition is to conserve genetic diversity. Species sub-structure is particularly important to anadromous salmonines because their strong homing capability fosters the formation of discrete populations (stocks) exhibiting important adaptations to local riverine ecosystems and the watersheds that determine their character (Berst and Simon 1981; Utter 1981; Utter et al. 1993; Nielsen 1998).In an effort to clarify the definition of species for Pacific salmon (Oncorhynchus sp.) under the ESA, Waples (1991a, b) proposed a more precise definition called the Evolutionarily Significant Unit (ESU). The purpose of this approach was to create a definition of species (DPS) for assessing Pacific salmon populations under consideration for ESA protection. The stock concept is the scientific foundation of the ESU approach. The stock concept has been subjected to critical peer-review and emphasizes the ecological importance of discrete populations in fish (Larkin 1981; Ricker 1981). Differences between stocks are important for management purposes since discrete stocks can vary in their productivity, population dynamics, and adaptations to local ecosystems (Ricker 1981; Hovey et al. 1989; Taylor 1991; Nielsen 1998; Verspoor 1997). The ESU approach facilitates a comprehensive analysis that categorizes units within a nested hierarchy ranging from a single stock level to subspecies. Stocks can be grouped into a comprehensive ESU unless there are clear evolutionarily important differences among stocks. Grouping of stocks does not, in and of itself, prevent river-specific management procedures necessary to protect sub-populations within the larger unit.
In February 1996, the Services published a policy to clarify their interpretation of the phrase "distinct population segment" for the purposes of listing, delisting and reclassifying species under the ESA (51 FR 4722). As related to salmonines, this policy encompasses the ESU policy and expands upon management issues related to jurisdictional issues. This joint NMFS-USFWS policy consists of three elements to be considered in a decision regarding the status of a possible DPS as endangered or threatened under the ESA: 1) the discreteness of the population segment in relation to the remainder of the species or subspecies to which it belongs; 2) the significance of the population segment to the species or subspecies to which it belongs; and 3) the conservation status of the population segment in relation to ESA listing standards. In this section of the status review, the BRT analysis focuses on the delineation of DPS structure for anadromous Atlantic salmon in U.S. Rivers and examines the first two elements - discreteness and significance. The conservation status of Atlantic salmon will be considered in subsequent sections of this status review in relation to the listing factors and efforts underway to protect the species (Section 7.1- 7.5).
Although the ESU approach was developed for Pacific salmon (Oncorhynchus sp.), Atlantic salmon populations have analogous population structure, ecology, and life history strategies. Throughout their range, Atlantic salmon are naturally substructured into genetically differentiated and reproductively isolated populations within and among drainages (Saunders 1981; Thorpe and Mitchell 1981; Stahl 1987; Bermingham et al. 1991; Nielsen 1998; King et al. 1993; 1999). Genetic exchange among stocks is minimized by spatial or temporal isolation during spawning. While straying occurs at low to moderate levels among populations of Atlantic salmon, it is typically not sufficient to prevent genetic divergence (Bermingham et al. 1991). Kendall (1935) first recognized that discrete Atlantic salmon stocks were found between U.S. rivers. Genetic and ecological data from other regions indicate that Atlantic salmon stocks are distinct at the river level and often between sections of larger drainages (Moller 1970; Verspoor et al. 1991; King et al. 1993; Nielsen et al. 1997; King et al. 1999). Given assessments of genetic stock structure and historic accounts of U.S. rivers, it is apparent that locally adapted, river-specific stocks of Atlantic salmon existed in the U.S., and that this structure was important to the overall fitness and productivity of the species (Kendall 1935; Riddell et al.1981; Baum 1997; Nielsen 1998; Thorpe et al. 1998). Furthermore, research on Atlantic salmon in Denmark has illustrated that even populations that have been decimated by anthropogenic factors can contain important genetic materials from ancestral stocks (Nielsen et al. 1997). Given these facts, the BRT concludes that population structure above the stock level is also a functional characteristic of Atlantic salmon stocks. To fully address the question of listing Atlantic salmon as a threatened or endangered species throughout its range, the BRT analyzed all relevant evidence of population structure within U.S. tributaries.
5.2 ANALYSIS OF DPS STRUCTURE WITHIN THE U.S.
The BRT examined genetic, life history, biogeographic, and environmental information in the process of evaluating Atlantic salmon throughout their U.S. range under provisions of the U.S. Endangered Species Act. The BRT found that distribution data, life history information, and ecoregion classification, were most useful for this process. Determining DPS structure is made more difficult by the fact that Atlantic salmon are absent from most (96%) of their historic freshwater rearing habitat in the USA. The Atlantic salmon in rivers south of the Kennebec River were extirpated by the mid-1800's to early 1900's (Atkins 1874; Kendall 1935) (Figure 4.1.1). Human activities moved the southern range of Atlantic salmon almost 2o north in latitude and eliminated the species from three of the largest Atlantic salmon rivers in the U.S: the Connecticut, Androscoggin, and Merrimack Rivers.Some populations north of the Kennebec River have been extirpated as well. These northern rivers were primarily small rivers with one hectare or less of available nursery habitat. The largest of these coastal rivers that lost their remnant stock was the Union River. It is unlikely that Atlantic salmon in all of these small watersheds were able to maintain persistent spawning runs given overall human perturbations of the past century. The small size of many these rivers and streams might have made these populations ephemeral (e.g. supported by low-level straying from larger neighboring populations), even under pre-colonial conditions. However, the unique genetic composition and persistence of the Atlantic salmon population of Cove Brook (Buckley 1999; King et al. 1999) indicates that retention of remnant stocks in these small rivers is possible. An examination by the BRT of the location of small rivers with documented juvenile Atlantic salmon juveniles suggests that the probability of a smaller drainage still having naturally producing populations is enhanced by close proximity to larger populations and overall habitat quality within the small drainage.
5.2.1 Extirpated Populations
Despite the limited historical information, assessing the probable historical structure of extirpated populations is important to categorizing the remaining populations and is informative in delineating their natural historical range. The BRT used the documented absence of wild Atlantic salmon from natal habitat for at least two generations (12 years) as the criterion for the total loss of a native population. We chose this conservative definition because the complex life history of Atlantic salmon often includes a total of six year classes (three freshwater and three marine) extant in the population at any one time. This life history adaptation is an important buffer against ecological disasters, both natural and human induced. We chose two generations to increase the certainty that failure to detect the presence of fish was a result of absence, not low numbers of fish and limited effort.Because little historical biological data is available for rivers with extirpated populations, it was necessary to use a zoogeographical approach within the context of Atlantic salmon ecology and historical range. To facilitate this analysis, the BRT utilized zoogeographical maps of boundaries between areas that would likely have different selective pressures for Atlantic salmon populations and substantial differences in riverine-marine ecosystem structure and function. Key elements to these determinations were: 1) the spatial arrangements of river systems that create isolation, and 2) watershed location within ecological provinces and subregions that affect the productivity and ecology of riverine-marine ecosystem complexes. Ecological provinces are areas of relatively uniform ecological potential that have been mapped based upon associations of environmental and biotic factors (Bailey 1995; Bailey 1998). These provinces are useful for modeling, strategic planning, and assessment because they synthesize the geoclimatic setting (climate, physical geography, water, soils, air, hydrology). All these geoclimatic factors influence river productivity and the structure and function of aquatic ecosystems and the transfer of energy in aquatic food chains (Vannote et al.1980; Cushing et al. 1983; Minshall et al. 1983; Cummins et al. 1984; Minshall et al. 1985; Waters 1993). The provinces that Bailey (1995) established have been delineated independent of watersheds. Incorporating aquatic ecological units (AEU) into analysis alleviates this weakness because AEUs reflect watershed linkage both to the sea and to other ecosystems. The defining criteria for an AEU in New England are the merging and separation of fish species assemblages caused by glaciation and orogeny (Maxwell et al. 1995). Particularly useful for DPS analysis are the subregions of the AEU. Subregions integrate major drainage systems within a region exhibiting similar endemic fish communities, accounting for historic mixing and isolation of fish populations. Using these ecological maps, the BRT determined that historic U.S. Atlantic salmon populations were minimally comprised of three DPSs that were primarily delineated by geography and juxtaposition.
5.2.1.1 Long Island Sound
A DPS is likely to have been present in the eight rivers that enter the Long Island Sound. These rivers are spatially isolated from the other Atlantic salmon rivers in New England by geographic features (Figure 4.1.1). Additionally, lower reaches of the Connecticut River and the entire watersheds of the others are characterized by the Eastern Broadleaf Forest Oceanic Province (Bailey 1995; Bailey 1998). A temperate deciduous forest dominates vegetation in this ecoregion that is between 0 and 300 m above sea level. Mean rainfall in this province is between 89 and 153 cm and frost free days range from 100 to 140. The upper Connecticut River watershed lies in the New England Mixed Forest Province at elevations above 300 m in regions dominated by mixed forests. Elevation has a substantial local influence on landscape ecology and watershed characteristics in these upper reaches. Most critically, all these systems enter Long Island Sound, a relatively large (175 km long, and 30 km wide) and shallow (24 m average depth) estuary. These tributaries and associated nearshore waters represent an AEU distinct from Gulf of Maine tributaries (Maxwell et al. 1995). All other Atlantic salmon rivers in the U. S. pass through river-dominated estuaries characterized by relatively narrow, short, cylindrical channels, before entering the Gulf of Maine. The more southerly position of Long Island Sound and its shallow nature provide substantially warmer nearshore waters than the Gulf of Maine. These substantial habitat differences, isolation, and interactions with different biotic communities than found in the Gulf of Maine, likely combined to make Atlantic salmon in this region unique. The Long Island Sound DPS was extirpated by the early 1800's, with the loss of Atlantic salmon stocks indigenous to the Connecticut River (Meyers 1994).5.2.1.2 Central New England
From Buzzards Bay northward along the coast of Massachusetts to the mouth of the Merrimack River, the BRT found no historical accounts of indigenous Atlantic salmon populations. This is likely a result of the small sizes and warm temperatures common to streams in this region. However, A DPS is likely to have existed in waters from the Merrimack River in the south to the Royal River (Yarmouth, Maine) in the north. Atlantic salmon populations in Central New England were probably distinct from those to the north because these populations south of Casco Bay are primarily in the Eastern Broadleaf Oceanic Province (EBOP) (Bailey 1995; Bailey 1998). The characteristic differences between the EBOP and the Laurentian Mixed Forest Province (LMFP) to the north, likely had a strong effect upon Atlantic salmon ecology and production. Elevations in Central New England portions of the EBOP range from sea level to 300 m, compared to the LMFP where elevations range up to 730 m. Additionally, the two largest rivers in this unit, the Merrimack River and the Saco River, extend into the Adirondack-New England Mountain Forest Province for approximately a third of their length. This creates a dramatic range in elevations throughout these drainages with a lower coastal tier to about 300 m rising to a mountainous habitat in the upper watersheds. Rainfall in the EBOP averages 89-153 cm, substantially higher than the 61-116 cm average in the LMFP. The transition from the EBOP to the LMFP is also characterized by a transition from deciduous forests to transitional conifer dominated forest types. All of these differences would influence the structure and function of aquatic ecosystems between these two provinces based on stream ecology theory (Vannote et al.1980; Cushing et al. 1983; Minshall et al. 1983; Cummins et al. 1984; Minshall et al. 1985; Waters 1993). Cumulatively, these differences likely made Atlantic salmon populations of Central New England distinct from those to the north. The Central New England DPS was extirpated in the mid-1800's, as documented by the loss of Merrimack River stocks (Stolte 1981, 1994). Early restoration efforts in the late 1800's had some success but the remnant populations of this region were likely extirpated before 1900 (Stolte 1981).5.2.1.3 Other Historic DPS Units
In addition to these broader DPSs above, larger rivers like the Connecticut, Merrimack, Androscoggin, Kennebec, and Penobscot likely contained unique stocks that may have contributed to historical DPS structure. These larger systems with unique riverine and lacustrine habitats are more heterogeneous than smaller river systems and this varied geo-physical nature likely fostered the development of unique stocks. Baum (1997) reports that Atlantic salmon in Maine rivers historically exhibited early-run and late-run components thought to be separate stocks. Additionally, it was thought that some of the larger, early-return fish on the Penobscot spawned in large river habitat in the upper reaches of the watershed. This account was based on the hypothesis that larger fish would be the only individuals capable of ascending some of the cascades, rapids, and falls in this river system. This evidence indicates that some of these larger rivers may have had populations that were quite different from coastal stocks and contained multiple DPSs within a river. It is possible that a specific component of a stock complex within a river was closely related to coastal populations while other fragments were quite different. Most of these large river populations, with the exception of the Penobscot, have been extirpated and documentation of ecological traits is scarce. Thus, it is difficult to determine the extent of the genetic resource in the U.S. that has been lost. The BRT's operating hypothesis that larger rivers may have contained unique DPS(s) is consistent with observations of differences between Pacific salmon stocks in coastal versus inland rivers classified in the Pacific Northwest (e.g., Southern Oregon/Northern California Coasts ESU and Lower Columbia River/Southwest Washington ESU of coho salmon). The majority of any unique large-river DPS(s) have been extirpated, with the possible exception of the Penobscot River stock that is in the process of being rebuilt through hatchery enhancement and supplementation (Table 4.2.2)(Baum 1997). Additionally, smaller streams, below the first major dams on lower Penobscot and Kennebec drainages, may have served as refugia for Atlantic salmon blocked from natal mainstem habitat. However, it is likely that decades of isolation from mainstem habitat subjected these populations to different selective pressures, altering their adaptability for large river systems. Thus, remnant populations in these smaller drainages are likely not simply relic populations from larger rivers but part of a larger population unit (DPS or restoration stock).The loss of naturally reproducing fish in the Connecticut and Merrimack Rivers represented nearly 40% of historic U.S. Atlantic salmon juvenile production habitat (Figure 4.1.2). The loss of these two southern-most rivers and their indigenous Atlantic salmon populations certainly had an influence on the genetic diversity of this species in the U.S. and North America. These rivers are currently the focus of restoration efforts using nonindigenous stocks mostly of Penobscot River origin. While restoration of Atlantic salmon in these rivers is beyond the scope of the ESA, their restoration would represent a significant contribution to the U.S. Atlantic salmon resource. Return rates from stocking in the Connecticut and Merrimack Rivers have been poor relative to other North American stocks (Saunders 1981; Friedland et al. 1993). Reasons for these low return rates appear to be attributable to the loss of local adaptations to unique habitat characteristics associated with the extirpated stocks (Saunders 1981). Other research supports this hypothesis and indicates that when stocks are transferred to new river systems, those from nearby rivers typically exhibit higher return rates than stocks from rivers farther away (Ritter 1975; Reisenbichler and McIntyre 1977; Riddell et al. 1981; Ritter et al. 1986; Hopley 1989). Additionally, stock specific differences in susceptibility to bacterial and viral diseases, underscore the importance of genetic variability not only to the viability of local stocks but as a genetic resource for conservation, restoration, and commercial aquaculture applications (Gjedrem and Gjoen 1995). The loss of locally adapted stocks has made restoration more difficult. Fortunately, salmonine populations show evidence of plasticity when introduced to new environments, and locally adapted and genetically differentiated stocks have developed in less than 20 generations (Krueger et al. 1994). As these restoration programs continue, their focus on the development of river-specific stocks should enhance the genetic resources of Atlantic salmon in the United States.
5.2.2 Extant Populations - Gulf of Maine DPS
The Gulf of Maine DPS includes all coastal watersheds with native populations of Atlantic salmon north of and including tributaries of the lower Kennebec River (below Edwards Dam) to the mouth of the St. Croix River at the US-Canada border. The DPS includes both early- and late-run Atlantic salmon (Baum 1997). Present low abundance levels and limited historical documentation preclude a quantitatively-based separation of these populations based upon run timing.Historically, the Androscoggin River delimited the DPS to the south, but populations south of the Kennebec River have been extirpated. The BRT delimited the geographical southern limit of the Gulf of Maine DPS as the southern border of the Laurentian Mixed Forest Province (LMFP)(Bailey 1995; Bailey 1998). The characteristic differences between the LMFP and the EBOP to the south likely had a strong effect upon Atlantic salmon ecology and production. Elevations in the LMFP are generally higher, ranging up to 730 m, but do not extend into mountain provinces; annual precipitation is lower, with differing seasonal regimes (Bailey 1995). These differences would influence the structure and function of aquatic ecosystems (Vannote et al.1980; Cushing et al. 1983; Minshall et al. 1983; Cummins et al. 1984; Minshall et al. 1985; Waters 1993) and create a different environment for the development of local adaptations than rivers to the south. Additional, zoogeographical rationales for the southern boundary were presented in the discussion of the Central New England DPS (Section 5.2.1.2).
The northern extent of this DPS likely included the St. Croix River and southwestern New Brunswick coastal tributaries northward to the St. John River. Inner-Fundy Atlantic salmon populations are very distinct from the Gulf of Maine DPS since they are primarily 1SW fish that do not migrate to the Labrador Sea (Marshall 1998). Available information also indicates that St. John River populations were historically dominated (greater than 50%) by 1SW Atlantic salmon; thus a portion of the population(s) was likely distinct from stocks to the south including the St. Croix River (Baum 1998; Marshall 1998). Additionally, the St. John River is another of the larger Atlantic salmon watersheds that may have contained stocks that were divergent from coastal river populations. The Canadian portion of this DPS is likely extirpated and under restoration. A possible exception is the St. John River population that contains a much larger 1SW component than any U.S. remnant or restoration stock but is largely derived from river-specific broodstock (Marshall 1998; Parrish et al. 1998). These Canadian rivers, including the mainstem St. Croix River, were eliminated from consideration because the joint NMFS-USFWS policy on DPS (61 FR 4722) facilitated separation by international management and exploitation differences for border stocks (62 FR 66325) (Section 5.3.2).
The Penobscot River was not included in the DPS at this time. This decision was based on the lack of a comprehensive genetic survey of the mainstem Penobscot River stock (hatchery and wild returns). Because potentially important and heritable adaptations are needed for larger river systems, it would be premature to determine the status of this population in relationship to the Gulf of Maine DPS without comprehensive genetic data. Sample collections, genetic analyses, and biological information are still being collected by the FWS and will analyzed to make a final determination of the status of the Penobscot River population relative to the coastal Atlantic salmon populations of the Gulf of Maine DPS. Collection of the 1999 samples will be complete in October of 1999 and analyses of these data should be completed by winter of this year. It is important to note that tributaries of the lower Penobscot estuary (south of the Bangor Dam) are considered within the DPS range.
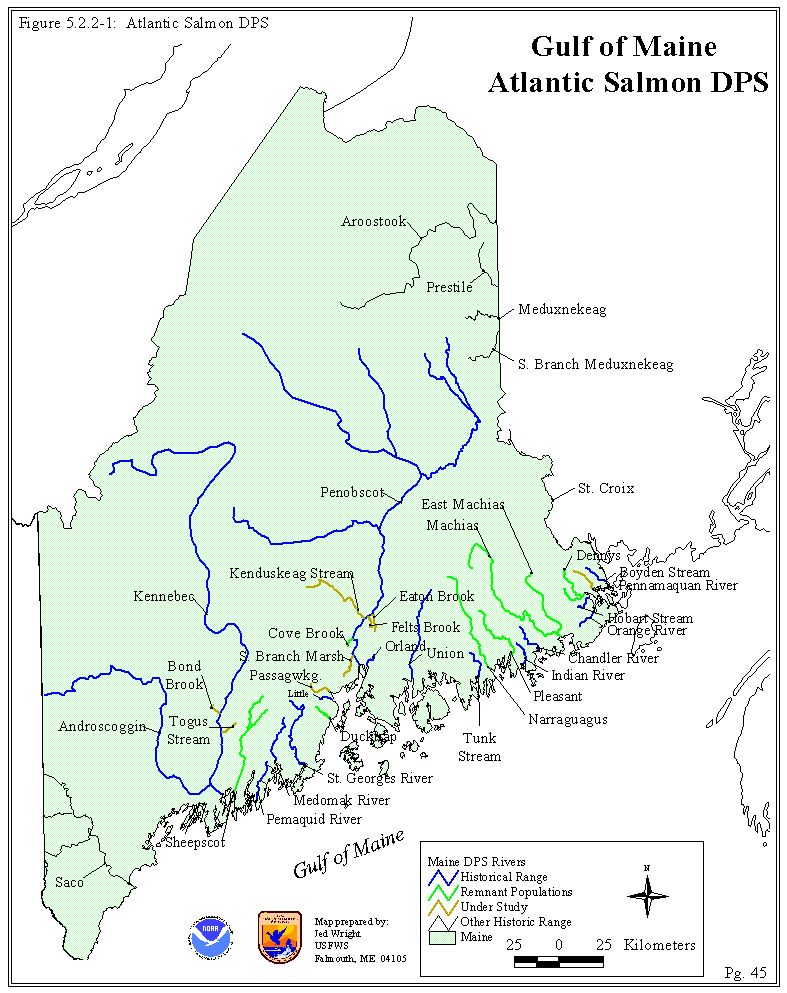
There are at least eight rivers in the DPS range that still contain functioning populations, albeit at substantially reduced abundance levels (Baum 1997; King et al. 1999). The core of these remnant populations is located in the Dennys, East Machias, Machias, Pleasant, Narraguagus, Ducktrap, and Sheepscot Rivers. These watersheds are among the largest of the historic river habitat currently accessible, averaging greater than 300,000 m2 of juvenile production habitat (Figure 4.2)(Baum 1997). The smallest of these seven populations is the Ducktrap River with 80,000 m2 of juvenile production habitat. Recent survey work also indicates that a naturally producing population that is genetically distinct (alleles only found in that population) remains in Cove Brook as well (Buckley 1999; King et al. 1999). This information demonstrates the ability of an Atlantic salmon population to retain unique genetic material in a relatively small drainage - juvenile habitat area in Cove Brook is estimated at only 23,500 m2 (Ed Baum, Atlantic Salmon Authority, personal communication). Additional surveys have also identified juvenile Atlantic salmon to be present in other river systems with relatively limited juvenile production habitat such as Bond, Togus, Passagassawaukeag, Eaton, Felts, South Branch Marsh, Kenduskaeg, and Pennamaquan Rivers (Buckley 1999)(Figure 5.2.2-1). Future routine fish surveying work and surveys initiated by public inquires may identify more populations that warrant study. The status of these populations relative to their demographic history and genetic legacy (DPS, aquaculture strays, Penobscot restoration stock strays, etc.) is being addressed by the Services on a case-by-case basis. Dams also block several of the drainages in within the historic DPS range, limiting Atlantic salmon recolonization (e.g., Medomak, St. George, and Union Rivers). Of these additional drainages, only the Union River is frequently stocked (Penobscot River origin fish). The other drainages are not targeted for stocking by Maine or Federal fisheries agencies although a few Atlantic salmon fry from educational programs in Maine schools may have been released in isolated cases. A detailed discussion of the factors that make these populations unique is included in Section 5.3.
Figure 5.5.2-1: Gulf of Maine Atlantic Salmon DPS
{kind=link}
5.3 ASSESSMENT OF THE GULF OF MAINE DPS
5.3.1 Species Status Elements of the Services' DPS Policy
The joint NMFS-USFWS policy (61 FR 4722) consists of two elements to be considered in a decision regarding the classification of potential vertebrate population segments as endangered or threatened under the ESA:- the discreteness of the population segment in relation to the remainder of the species or subspecies to which it belongs, and
- the significance of the population segment to the species or subspecies to which it belongs. Subsections 5.3.2 - 5.3.4 address these two elements.
5.3.2 Discreteness
According to the Services' DPS policy (61 FR 4722), a population segment may be considered discrete if it satisfies either of the following two conditions:- it is markedly separated from other populations of the same taxon as a consequence of physical, physiological, ecological or behavioral factors; or
- it is delimited by international governmental boundaries across which there is a significant difference in control of exploitation, management of habitat, or conservation status.
5.3.2.1 Separateness From Other Populations
Reproductive isolation does not have to be absolute to allow evolutionarily important differences to accrue in different population units, only strong enough for these differences to develop and be maintained (Wright 1978; Waples 1991). Geographical distance, behavioral differences, and/or temporal segregation of spawners can maintain reproductive isolation. The occurrence of nonindigenous Atlantic salmon in a stream does not necessarily represent a breakdown of reproductive isolation unless these fish spawn successfully, their progeny survive to spawn, and their presence degrades the survival and fitness of native stocks. In fact, some genetic exchange between populations helps to maintain genetic fitness by countering genetic drift (Waples 1991). To examine whether Gulf of Maine populations are separate from other populations, the BRT examined three major indicators: straying of spawners; recolonization rates external to the DPS; and genetic differences observed throughout the range of Atlantic salmon.The Gulf of Maine DPS of Atlantic salmon is markedly separated from other populations of the same taxon. The zoogeographical basis of this separation within the U.S. is described in detail in Section 5.2. All populations south of this DPS have been extirpated. This separation is the consequence of strong fidelity to natal rivers and adaptations for local conditions that are well documented for all anadromous salmonines (Staahl 1981; Utter 1981; Utter et al. 1993b).
Comprehensive tagging and recovery experiments have been conducted in Maine Atlantic salmon rivers. Baum and Spencer (1990) assessed the homing of 1.2 million Carlin tagged Atlantic salmon stocked as smolts from 1966-1987 in five Maine rivers. Only 2% of the tag recoveries (n= 3,755) were from non-natal (stocking location) rivers. In addition, some fish recorded in one river as strays eventually returned to their natal stream, which indicates that weir and trap recaptures may overestimate the number of fish that actually spawn in non-natal rivers (Baum 1997). Data for discrete wild Atlantic salmon populations in Norway show straying rates ranging from 5% to 8% (Hansen and Jonsson 1994). Other researchers have shown that the straying rate of wild fish is typically lower than that of hatchery fish (Stabell 1984; Piggins 1987; Jonsson et al. 1991). Available information indicates that U.S. Atlantic salmon stocks do not stray far from their natal stream and thus supports the hypothesis that most straying documented between these river systems is limiting to neighboring rivers within the DPS geographical region. As such, the 2% straying rates documented by Baum and Spencer (1990) indicates that interactions between populations in individual DPS rivers and the Penobscot River restoration program are no greater than would be expected in wild populations of similar geographic proximity. The information in the tagging database indicates that specific stocks have persisted in several U.S. river systems and comprise the component parts of the Gulf of Maine Atlantic salmon DPS.
Isolation typically leads to genetic differences among Atlantic salmon stocks and relatively minor genetic differences have been linked to important adaptive morphological and life-history traits (Ritter 1975; Saunders 1981). These traits are important to the survival of individual stocks, DPSs, and the species as a whole. Atlantic salmon populations in Britain and Wales were found to be distinct using mitochondrial DNA analysis; additionally, differences in survival and migration were identified using shifts in clonal frequencies (King et al. 1993). Stahl (1987) stated that "Atlantic salmon are naturally substructured into multiple genetically differentiated and more or less reproductively isolated stocks within as well as between drainages." Given the existence of distinct stocks, creating artificially large gene flow between previously reproductively isolated populations would likely disrupt locally adapted gene complexes and decrease the overall productivity of the species (Stahl 1987).
North American Atlantic salmon stocks have been found to be distinct from European stocks using both electrophoretic and mitochondrial DNA analyses (Stahl 1987; Bermingham et al. 1991; Taggart et al. 1996). Recent data from King et al. (1999) further supports these differences and has provided analytical methods to distinguish continent-of-origin with 100% accuracy. In all these studies, genetic differences are strongly geographically patterned and, while variation is low compared to freshwater fish, it is consistent with results from other anadromous species (King et al. 1999). The genetic differences between North American and European Atlantic salmon are substantial enough that introgression of these stocks is likely to be detrimental (King et al. 1999).
Genetic differences have previously been found between U.S. and Canadian Atlantic salmon stocks using electrophoretic and DNA fingerprinting methods (Moller 1970; Bentzen and Wright 1992). In the most comprehensive survey to date, King et al. (1999) found that while genetic differences between U.S. and Canadian stocks sampled were small, assignment tests correctly classified U.S. fish to U.S. rivers more than 77% of the time. Canadian fish were correctly classified to Canadian rivers over 93% of the time. An additional 6% of the misclassified U.S. fish were classified to Canadian rivers in neighboring New Brunswick and Nova Scotia (i.e., 83% within a narrow geographical area). These data indicate that observed genetic differences between U.S. and Canadian Atlantic salmon stocks might represent important population differentiation. This differentiation is reinforced by the spatial distribution of Atlantic salmon rivers and differences in life history. Differences in life history are best illustrated by examination of the Bay of Fundy Atlantic salmon stocks. These stocks do not undertake extensive ocean migrations, remaining instead in the Bay of Fundy for most of the marine phase of their life cycle that is only 1SW (Marshall 1998). Populations in the St. Croix River and the St. John River, have intermediate life histories with a larger 1SW component than is found in any U.S. stock (Baum 1997). Since age-at-maturity appears to be a trait that is least partially genetically determined (Glebe and Saunders 1986; Ritter et al. 1986; Saunders 1986), this provides further evidence of separateness of U.S. and Canadian Atlantic salmon populations.
Some authors have claimed that the magnitude of past stocking efforts has facilitated introgression and eliminated local variability (Kornfield et al. 1995). While the historic isolation of stocks within the DPS may have been greater and supported higher levels of genetic differences, subtle distinctions between stocks within the DPS remain and differences from populations outside the DPS are clear (King et al. 1999; section 5.3.2.1.1). In particular, different methods have illustrated unique genetic material in two rivers, the Ducktrap River and Cove Brook (Kornfield 1994; King et al. 1999). The majority of stocks utilized for supplemental stocking within the Gulf of Maine DPS have been from within the DPS geographic range in both the USA and Canada (Baum 1997). Because the source of most stocking efforts has been from within the DPS, the genetic effects from stock mixing would be substantially less than from stocks from outside the DPS. These facts indicate that any inter-stock mixing that resulted from hatchery practices could have potentially elevated natural levels of intra-DPS genetic exchange beyond levels observed in the absence of stocking. An alternative hypothesis would be that the low levels of genetic differentiation between stocks is a natural consequence of a historic population structure characterized by low-to-moderate levels of genetic exchange between individual stocks in a metapopulation. Additionally, because most genetic analyses focus on neutral characters, they do not provide a complete and conclusive measure of distinctness or ecological adaptation. Failure to find a significant genetic difference between stocks may be the result of technological and/or sampling limitations and does not necessarily mean that differences with a genetic basis do not exist. The analysis of genetics data only provides some of the evidence for the separateness of DPSs.
The separateness of coastal stocks of the Gulf of Maine DPS and other Atlantic salmon populations outside the DPS are strongly supported by the persistence of these populations, geographic segregation, limited opportunities for stocking from outside the DPS to cause introgression, and current genetic analyses. The BRT concludes that adequate genetic and demographic data demonstrate that an ecologically important separation exists between the Gulf of Maine DPS within its current range and other populations to the north; all naturally occurring populations south of the DPS have been extirpated.
5.3.2.1.1 Intra-DPS Separateness
The examination of the separateness between populations within the DPS is of importance to the conservation of genetic materials within the DPS and ongoing management practices. A discussion of these differences is included here to document the intra-DPS genetic variability and the importance of conserving the component populations of the DPS. Historical accounts indicate that differences existed among river-specific stocks of Atlantic salmon. Kendall (1935) stated that in their natural state, U.S. rivers were "frequented by a sufficient number of salmon each year... and not enough migrants from other birth places entered them to prevent the establishment of somewhat differing races, peculiar to their respective streams." Recent studies examining the genetic differences among U.S. Atlantic salmon stocks have yielded somewhat contradictory and inconclusive results (Roberts 1976; Bentzen and Wright 1992; King and Smith 1994; Kornfield 1994; May et al. 1993, 1994; Schill and Walker 1994; King et al. 1999). Some researchers have found differences between select U.S. Atlantic salmon populations including unique alleles (Bentzen and Wright 1992; Kornfield 1994; King et al. 1999). Other researchers have not been able to demonstrate substantial differences between some of these stocks using different methods (Roberts 1976; King and Smith 1994; May et al. 1994; Schill and Walker 1994). These results are not unexpected given that different techniques often provide differing results (Park and Moran 1994). Collectively these studies indicate that there are differences between populations in the Gulf of Maine DPS and some of these populations contain unique genetic material.Gene flow between Gulf of Maine populations were estimated by Schill and Walker (1994), who surveyed five random amplified polymorphic DNA (RAPD) markers in nine U.S. Atlantic salmon populations. Their data suggest that a high rate of genetic flow occurs among the Atlantic salmon populations surveyed (2.9 to 4.1 effective migrants per generation). Likewise, May et al. (1994) measured gene flow on the order of 7.1 effective migrants per generation. These results are similar to those reported by King et al. (1999) in their more comprehensive analysis of molecular variance tests of 11 Maine rivers. These data indicate that some gene flow occurs between the populations that are part of the DPS but it does not preclude the maintenance of adaptive differences (May et al. 1994; Schill and Walker 1994; King et al. 1999). Genetic differentiation between stocks can still persist with low levels of gene flow and this gene flow helps to counter (non-adaptive) founder effect changes (Wright 1978; Waples 1991b). While, gene flow can act to inter-mix neutral alleles, selection for locally adapted alleles can counter homogenization and may strengthen the fitness of a population (Slatkin 1987). Since the values reported by King et al. (1999) measure both past and present gene flow between these populations, they may be somewhat elevated due to artificial mixing of stocks through past stocking practices (Section 4: Table 4.1) in this region. However, the temporal stability of allele frequencies and the occurrence of alleles that are unique to Maine indicate that this mixing has not overwhelmed all of the genetic differences between stocks (King et al. 1993; King et al. 1999).
The BRT concludes that while it is unlikely that any U.S. Atlantic salmon populations exist in a genetically pure native form, their continued presence in indigenous habitat indicates that important heritable local adaptations likely still exist. The conservation of the populations within the Gulf of Maine DPS is essential because these Atlantic salmon represent the remaining genetic legacy of ancestral populations that were locally adapted to the rivers and streams of the region. Some disagreements exist in the management of river-specific conservation programs that warrant continued debate (e.g. Kornfield et al. 1995 and Moring et al. 1995)
5.3.2.2 International Boundaries
The Gulf of Maine DPS represents the remaining genetic legacy of a U.S. Atlantic salmon resource that formerly extended from the Housatonic River to the headwaters of the Aroostook River. The northern range of the Gulf of Maine DPS is delimited not only by the natural zoogeographical constraints on local adaptations but by an international boundary. There are substantial differences in the control of exploitation, management of habitat, conservation status, and regulatory mechanisms of Atlantic salmon between the U.S. and Canada (May 1993; Baum 1997). Management and conservation programs in the United States and Canada have similar goals, but differences in legislation and policy support the use of the United States/Canada international boundary as a measure of discreteness for the purposes of evaluating stock status (NASCO 1999). Highlighting these differences is the lack of aboriginal or commercial fisheries for Atlantic salmon in the marine waters of the U.S. (Baum 1997; USASAC 1999). Additional factors that are significant relative to section 4(a)(1)(D) of the ESA are differences in regulatory mechanisms between the federal and state system that exists in the U.S. and the federal and Provincial governments in Canada.5.3.2.3 Evaluation of Discreteness
Based on the information available, the BRT concluded that the Gulf of Maine DPS of Atlantic salmon meets both criteria for discreteness as outlined in the Services' Policy on DPS evaluation. Only one of these is needed to conclude that the DPS is discrete from other populations.5.3.3 EVOLUTIONARY AND ECOLOGICAL SIGNIFICANCE
The second element of the Services' policy is the consideration of the population segment's biological and ecological significance to the taxon to which it belongs. This consideration may include, but is not limited to, the following: persistence of the discrete population segment in an ecological setting unusual or unique for the taxon; evidence that the loss of the discrete population segment would result in a significant gap in the range of a taxon; evidence that the discrete population segment represents the only surviving natural occurrence of a taxon that may be more abundant elsewhere as an introduced population outside its historic range; or evidence that the discrete population segment differs markedly from other populations of the species in its genetic characteristics.5.3.3.1 Persistence and Habitat Characteristics
Riverine habitat occupied by the Gulf of Maine DPS of Atlantic salmon is unique in that it is at the southern extent of the North American range of Atlantic salmon (Saunders 1981; Baum 1997). This habitat is also distinct because it includes core Atlantic salmon populations within a unique ecoregion, the Laurentian Mixed Forest Province of coastal Maine (Bailey 1995; Bailey 1998). With their location at the southern extent of the range, riverine habitat in the DPS rivers provides a more productive juvenile nursery habitat than most Canadian river systems, allowing U.S. rivers to produce proportionately more one and two-year smolts (Meyers et al.1986; Baum 1997; Hutchings and Jones 1998). Despite habitat differences in freshwater range, both U.S. and Canadian Atlantic salmon spend much of their marine life in the northwest Atlantic Ocean. By sharing feeding areas with other North American stocks and migrating from the most southern spawning areas, U.S. salmon undertake the longest oceanic migrations of the species in North America. Occupation of the southern portion of the range exposes U.S. salmon to riverine and oceanic selection factors different from those experienced by more northern stocks.Within the U.S., Atlantic salmon historically occurred in two general types of drainages: short (<125 km long) coastal rivers with few tributaries and relatively large river systems (>125 km long with most >160 km) with numerous tributaries. The remnant populations of the DPS represent populations from short coastal rivers. The Canadian segment of the historical DPS would have only contained two rivers of similar size to core US populations, the Digdeguash and Magaduavic Rivers. These rivers have significantly higher 1SW returns than U.S. stocks (Marshall 1998), making them more representative of Canadian stocks. Other physical differences also exist among rivers within the DPS and between the DPS and other populations, ranging from soil types, the presence of headwater and mainstem lakes, gradient, and other physiochemical and geographic characteristics. Substantial environmental differences between watersheds typically foster the independent evolution and selection of locally adaptive heritable traits in salmonines (Stahl 1987; Utter 1981; Verspoor 1988; Claytor and Verspoor 1991; King et al. 1993). These processes would have influenced the evolution of U.S. Atlantic salmon stocks into separate populations within discrete DPSs.
To survive at the extreme southern range of the species, U.S. Atlantic salmon populations needed to be adapted to distinct physical and environmental challenges (Saunders 1981). These adaptations contribute to the distinctness of the Gulf of Maine DPS as an important population segment and make its survival important to the species as whole. The BRT determined that a critical factor in determining the status of U.S. Atlantic salmon stocks was the historic persistence of a naturally reproducing legacy stock in each component river. Atkins (1874), Kendall (1894;1935), Rounsefull and Bond (1949), Baum (1997), and USASAC (1999) documented that these stocks as a collective group, the Gulf of Maine DPS, have persisted over time. While some citations suggest localized extinction of individual populations, the basis for this is somewhat speculative given rapid population growth when an impoundment was breached (e.g. Rounsefell and Bond 1949). Additionally, the recolonization of individual stocks appears to be primarily driven by natural processes of recolonization from within river (below impoundment) and within DPS (neighboring river) refugia. The success of stocking efforts prior to 1971 has been evaluated as being extremely poor (Fletcher 1955; Baum 1997). The failure of these early stocking programs was likely a result of the biotechnological inefficiency of early hatchery practices (Baum 1997) and the use of nonindigenous stocks (Saunders 1981). Most stocking efforts since 1970 have used U.S. origin fish (Penobscot River and Gulf of Maine DPS) and all stocking within the Gulf of Maine DPS since 1991 has been river-specific in origin (Baum 1997; USASAC 1999). The overall average run composition from 1970 to present exceeds 80% wild-origin for the seven rivers monitored (USASAC 1999)(Section 5.3.3.4). The balance of hatchery returns in the last 25 years is comprised almost exclusively of Penobscot hatchery origin fish. A single Miramichi River grilse that ascended the Penobscot River in 1974 is the only documented return of a Canadian-origin fish from outside the Gulf of Maine DPS since 1960 (Baum 1997).
The BRT concludes that there is substantial evidence that remnant populations of the Gulf of Maine DPS have persisted in their native range. The unique geographical location and inherent ecoregion differences led the BRT to conclude that the Gulf of Maine DPS occupies an ecological setting that is unique for Atlantic salmon. The loss of this DPS would result in a significant gap in the range of this taxon, moving the range of this population an additional degree of latitude to the north. The loss of these populations would restrict the natural range of Atlantic salmon above the 45th parallel and beyond the borders of the U.S.A.
3.2 Phenotypic Traits
It is difficult to assess the importance of phenotypic characteristics for Atlantic salmon since temporal variation within stocks is extensive (Blouw et al. 1988; Hutchings and Jones 1998). However, Claytor and Verspoor (1991) found meristic and morphometric differences among stocks of Atlantic salmon to be related to environmental variables on a clinal scale in both North America and Europe. The differences among stocks appeared to represent adaptations to environmental conditions that Atlantic salmon encounter throughout their range. Recent studies on phenotypic traits of U.S. Atlantic salmon have focused on juvenile fish. Morphometric and meristic variation indicated that U.S. populations possessed a large number of traits that differed from the population of the St. John River in Canada (Kincaid et al. 1994). Variation in measurements between the seven U.S. rivers surveyed indicated that the potential for stock-specific differences existed for some traits. These data indicate that phenotypic traits are unique among these geographic locations. Atkins (1874) noted historical differences in the size of mature, 2SW adults, among U.S. rivers. In addition, differences in the shape and girth of Atlantic salmon were reported in some rivers (Kendall 1935). The recent studies and historical accounts of phenotypic traits listed above indicate that stock-specific differences existed historically and are now present.3.3 Life History Characteristics
Differences in life history among U.S. Atlantic salmon stocks and those of Canada were identified as early as 1874 (Atkins 1874). U.S. Atlantic salmon stocks have been composed of predominately 2SW salmon (> 80%) from at least the late 1800's to the present (Atkins 1874; Kendall 1935; USASAC 1999). Alternately, many Canadian stocks and several in Europe have a much higher grilse component with a concurrently lower 2SW component that is frequently less than 50% (Hutchings and Jones 1998). This life history trait is partially controlled by stock genetics (Ritter et al. 1986). Because U.S. stocks have return age composition that differs from Canadian stocks, especially neighboring stocks in the Scotia-Fundy region, it can be inferred that the genetic component of this trait also differs. The predominance of 2SW fish also influences spawning-run timing because they typically enter rivers earlier than grilse. Trends in run timing are difficult to discern due to low abundance and the lack of collection facilities on all rivers, but analyses of the recreational catch in some Maine rivers has indicated that the timing of spawning runs has changed little in the past 50 years (Baum 1997). Atkins (1874) also hypothesized that differences in smolt emigration timing occurred between rivers. Data for recent smolt migration trends are not available. Recent syntheses of juvenile Atlantic salmon data suggest that while environment has a strong influence upon juvenile growth and maturation (precocious parr), heritable differences between stocks also influence growth and performance (Kincaid 1994; Hutching and Jones 1998).Taking into account all of the foregoing factors, the BRT determined that differences in life history characteristics historically contributed to the distinctness of the Gulf of Maine DPS. Remnant stocks have maintained the most characteristic of these factors: smoltification at a mean age of 2 and predominant adult returns as 2SW fish (age 4). Since the proportion of 2SW fish in an Atlantic salmon stock has a documented genetic basis (Glebe and Saunders1986; Ritter et al. 1986; Hutchings and Jones 1998), the BRT concludes that the DPS has unique life history characteristics that have a heritable basis. The BRT concludes that both environmental and genetic factors make the Gulf of Maine DPS markedly different from other populations of Atlantic salmon in their life history and ecology.